V(D)J rekombinace je mechanismus genetické rekombinace u obratlovců, který náhodně vybírá a spojuje segmenty genů kódujících specifické proteiny zásadní pro fungování imunitního systému. Tento proces rekombinace dává vzniknout rozmanitému repertoáru molekul receptorů T buněk (TCR) a imunoglobulinů, nezbytných k rozpoznávání množství antigenů, ať už pocházejících z cizorodých bakterií, virů, parazitů, nebo vlastních poškozených buněk, zejména nádorových.
Struktura imunoglobulinů a receptorů T buněk
Lidské protilátky (a receptory B buněk) sestávají z těžkých (heavy) a lehkých (light) řetězců s konstantními (C) a variabilními (V) oblastmi. Jednotlivé řetězce jsou kódovány třemi typy genů:
- Gen kódující těžký řetězec se nachází na 14. chromosomu
- Gen kódující lehký řetězec kappa (κ) se nachází na 2. chromosomu
- Gen kódující lehký řetězec (λ) se nachází na 22. chromosomu
Množství genů kódujících variabilní oblasti každého z řetězců je seskupeno do tří segmentů. Lokus kódující těžký řetězec u člověka například obsahuje 65 genů ve V (variable) segmentu, 27 v D (diversity) segmentu a 6 v J (joining) segmentu. Pro lehké řetězce také existuje mnoho genů V a J, geny D zcela chybí. Mechanismem přesmyku DNA v těchto segmentech lze dosáhnout enormního repertoáru protilátek — 65 V × 27 D × 6 J genů = 10 530 kombinací jen u těžkých řetězců, počet možností ještě mnohonásobně zvyšuje obdobná variabilita lehkých řetězců.
Většina receptorů T buněk sestává z α (alfa) a β (beta) řetězce. Geny pro tyto receptory, podobně jako pro imunogobuliny rovněž obsahují V, D a J segmenty v β řetězcích (a V a J segmenty v α řetězcích), které se během vývoje T-lymfocytů stejným způsobem přeskupují a dávají tak vzniknout buňce s jedinečným povrchovým receptorem.
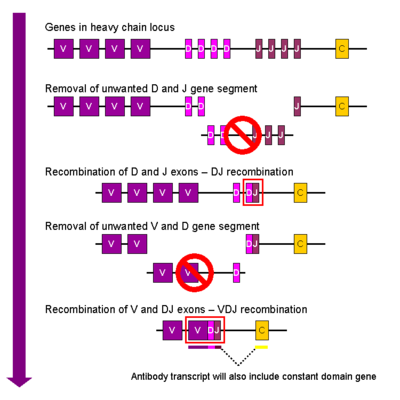
V(D)J rekombinace imunoglobulinů
Během vývoje B buněk dochází k první rekombinaci mezi jedním z D genů a jedním z J genů v lokusu pro těžký řetězec. Veškerá DNA mezi těmito geny je z genomu nenávratně vystřižena a odstraněna. Po této D-J rekombinaci následuje podobným způsobem připojení jednoho z V genů ke vzniklému DJ komplexu, čímž vzniká gen VDJ; veškeré geny mezi vybranými V a D geny jsou opět trvale odstraněny. Primární transkript (tj. nesestřižená RNA, dříve, než dochází ke splicingu) vždy obsahuje VDJ oblast a oba konstantní řetězce mí a delta (Cμ a Cδ; primární transkript tedy obsahuje segmenty V-D-J-Cμ-Cδ). Tato pre-mRNA je posttranskripčně modifikována přidáním poly (A) konce na 3′-konec mRNA a vystřižením sekvence mezi VDJ segmentem a Cμřetězcem. Translace této mRNA pak vede k produkci těžkého řetězce imunoglobulinu M.
Lokusy pro kappa (κ) a lambda (λ) řetězce imunoglobulinů se přeskupují podobným způsobem, chybí zde však D segment. V prvním kroku rekombinace zde tedy dochází k vytvoření VJ komplexu, při transkripci pak k připojení genu pro konstantní oblast. Translace sestřižené mRNA kódující κ nebo λ řetězce pak vytváří samotný Igκ nebo Igλ protein.
Spojením těžkého řetězce Igμ s jedním z lehkých řetězců vzniká membránová forma imunoglobulinu IgM (tzv. B-buněčný receptor), který je exprimován na povrchu nezralých B buněk.
V(D)J rekombinace receptorů T buněk
Během vývoje T buněk dochází v genech pro receptory T buněk (TCR, T cell receptor) ke stejným procesům, které byly výše popsány u imunoglobulinů. D-J rekombinace nastává nejdříve v genu pro β řetězec TCR, kde se připojuje buď gen Dβ1 k jednomu ze šesti Jβ1 genů, nebo gen Dβ2 k jednomu ze šesti Jβ2 segmentů. Po D-J rekombinaci následuje Vβ-DβJβ přesmyk. Všechny geny mezi příslušnými Vβ-Dβ-Jβ geny jsou nenávratně odstraněny. Na konec primárního transkriptu se vedle zmíněného VβDβJβ řetězce začleňuje gen pro konstantní oblast proteinu, Cβ. RNA splicing vystřihne všechny nadbytečné sekvence a hotovou mRNA lze posléze přeložit do formy Cβ řetězce TCR.
Přeskupení α řetězce TCR je analogické přeskupení β řetězce a podobá se V-J rekombinaci lehkých řetězců imunoglobulinů (viz výše). Spojení α a β řetězců dává vzniknout αβ-TCR, které na svém povrchu exprimuje většina T buněk.






